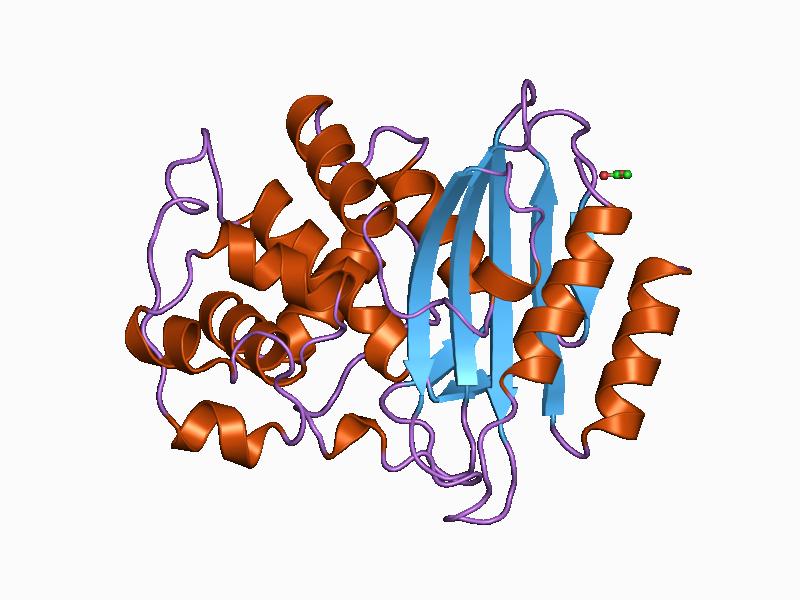
Beta-lactamase
Beta-lactamases (β-lactamases) are enzymes (EC 3.5.2.6) produced by bacteria that provide multi-resistance to beta-lactam antibiotics such as penicillins, cephalosporins, cephamycins, monobactams and carbapenems (ertapenem), although carbapenems are relatively resistant to beta-lactamase. Beta-lactamase provides antibiotic resistance by breaking the antibiotics' structure. These antibiotics all have a common element in their molecular structure: a four-atom ring known as a beta-lactam (β-lactam) ring. Through hydrolysis, the enzyme lactamase breaks the β-lactam ring open, deactivating the molecule's antibacterial properties.
Serine beta-lactamase
β-lactamase domain
?
Beta-lactamases produced by Gram-negative bacteria are usually secreted, especially when antibiotics are present in the environment.[1]
Structure[edit]
The structure of a Streptomyces serine β-lactamase (SBLs) is given by 1BSG. The alpha-beta fold (InterPro: IPR012338) resembles that of a DD-transpeptidase, from which the enzyme is thought to have evolved. β-lactam antibiotics bind to DD-transpeptidases to inhibit bacterial cell wall biosynthesis. Serine β-lactamases are grouped by sequence similarity into types A, C, and D.
The other type of beta-lactamase is of the metallo type ("type B"). Metallo-beta-lactamases (MBLs) need metal ion(s) (1 or 2 Zn2+ ions[2]) on their active site for their catalytic activities.[3] The structure of the New Delhi metallo-beta-lactamase 1 is given by 6C89. It resembles a RNase Z, from which it is thought to have evolved.
Mechanism of action[edit]
The two types of beta-lactamases work on the basis of the two basic mechanisms of opening the β-lactam ring.[2]
The SBLs are similar in structure and mechanistically to the β-lactam target penicillin-binding proteins (PBPs) which are necessary for cell wall building and modifying. SBLs and PBPs both covalently change an active site serine residue. The difference between the PBPs and SBLs is that the latter generates free enzyme and inactive antibiotic by the very quick hydrolysis of the acyl-enzyme intermediate.
The MBLs use the Zn2+ ions to activate a binding site water molecule for the hydrolysis of the β-lactam ring. Zinc chelators have recently been investigated as metallo-β-lactamase inhibitors, as they are often able to restore carbapenem susceptibility.[4]
Penicillinase[edit]
Penicillinase is a specific type of β-lactamase, showing specificity for penicillins, again by hydrolysing the β-lactam ring. Molecular weights of the various penicillinases tend to cluster near 50 kilodaltons.
Penicillinase was the first β-lactamase to be identified. It was first isolated by Abraham and Chain in 1940 from E. coli (which are Gram-negative) even before penicillin entered clinical use,[5] but penicillinase production quickly spread to bacteria that previously did not produce it or produced it only rarely. Penicillinase-resistant beta-lactams such as methicillin were developed, but there is now widespread resistance to even these.
Treatment of ESBL/AmpC/carbapenemases[edit]
General overview[edit]
In general, an isolate is suspected to be an ESBL producer when it shows in vitro susceptibility to the cephamycins (cefoxitin, cefotetan) but resistance to the third-generation cephalosporins and to aztreonam. Moreover, one should suspect these strains when treatment with these agents for Gram-negative infections fails despite reported in vitro susceptibility. Once an ESBL-producing strain is detected, the laboratory should report it as "resistant" to all penicillins, cephalosporins, and aztreonam, even if it is tested (in vitro) as susceptible. Associated resistance to aminoglycosides and trimethoprim-sulfamethoxazole, as well as high frequency of co-existence of fluoroquinolone resistance, creates problems. Beta-lactamase inhibitors such as clavulanate, sulbactam, and tazobactam in vitro inhibit most ESBLs, but the clinical effectiveness of beta-lactam/beta-lactamase inhibitor combinations cannot be relied on consistently for therapy. Cephamycins (cefoxitin and cefotetan) are not hydrolyzed by majority of ESBLs, but are hydrolyzed by associated AmpC-type β-lactamase. Also, β-lactam/β-lactamase inhibitor combinations may not be effective against organisms that produce AmpC-type β-lactamase. Sometimes these strains decrease the expression of outer membrane proteins, rendering them resistant to cephamycins. In vivo studies have yielded mixed results against ESBL-producing K. pneumoniae. (Cefepime, a fourth-generation cephalosporin, has demonstrated in vitro stability in the presence of many ESBL/AmpC strains.) Currently, carbapenems are, in general, regarded as the preferred agent for treatment of infections due to ESBL-producing organisms. Carbapenems are resistant to ESBL-mediated hydrolysis and exhibit excellent in vitro activity against strains of Enterobacteriaceae expressing ESBLs.
Use as a pharmaceutical[edit]
In 1957, amid concern about allergic reactions to penicillin-containing antibiotics, a beta-lactamase was sold as an antidote under the brand name neutrapen.[36] It was theorized that the breakdown of penicillin by the enzyme would treat the allergic reaction.[37] While it was not useful in acute anaphylactic shock, it showed positive results in cases of urticaria and joint pain suspected to be caused by penicillin allergy.[38][39] Its use was proposed in pediatric cases where penicillin allergy was discovered upon administration of the polio vaccine, which used penicillin as a preservative.[40] However, some patients developed allergies to neutrapen.[41][42] The Albany Hospital removed it from its formulary in 1960, only two years after adding it, citing lack of use.[43] Some researchers continued to use it in experiments on penicillin resistance as late as 1972.[44] It was voluntarily withdrawn from the American market by 3M Pharmaceuticals in 1997.[45]
Detection[edit]
Beta-lactamase enzymatic activity can be detected using nitrocefin, a chromogenic cephalosporin substrate which changes color from yellow to red upon beta-lactamase mediated hydrolysis.[46]
Extended spectrum beta lactamase (ESBL) screening can be performed using disk-diffusion. Cefpodoxime, ceftazidime, aztreonam, cefotaxime, and/or ceftriaxone discs are used.[47]
Evolution[edit]
Beta-lactamases are ancient bacterial enzymes. Metallo β-lactamases ("class B") are all structurally similar to RNase Z and may have evolved from it. Of the three subclasses B1, B2, and B3, B1 and B2 are theorized to have evolved about one billion years ago, while B3 seems to have arisen independently, possibly before the divergence of the Gram-positive and Gram-negative eubacteria about two billion years ago.[48] PNGM-1 (Papua New Guinea Metallo-β-lactamase-1) has both metallo-β-lactamase (MBL) and tRNase Z activities, suggesting that PNGM-1 is thought to have evolved from a tRNase Z, and that the B3 MBL activity of PNGM-1 is a promiscuous activity and subclass B3 MBLs are thought to have evolved through PNGM-1 activity.[49] Subclasses B1 and B3 has been further subdivided.[50]
Serine beta-lactamases (classes A, C, and D) appear to have evolved from DD-transpeptidases, which are penicillin-binding proteins involved in cell wall biosynthesis, and as such are one of the main targets of beta-lactam antibiotics.[51] These three classes show undetectable sequence similarity with each other, but can still be compared using structural homology. Groups A and D are sister taxa and group C diverged before A and D.[52] These serine-based enzymes, like the group B betalactamases, are of ancient origin and are theorized to have evolved about two billion years ago.[53]
The OXA group (in class D) in particular is theorized to have evolved on chromosomes and moved to plasmids on at least two separate occasions.[54]
Etymology[edit]
The "β" (beta) refers to the nitrogen's position on the second carbon in the ring. Lactam is a blend of lactone (from the Latin lactis, milk, since lactic acid was isolated from soured milk) and amide. The suffix -ase, indicating an enzyme, is derived from diastase (from the Greek diastasis, "separation"), the first enzyme discovered in 1833 by Payen and Persoz.[55]