Gene flow
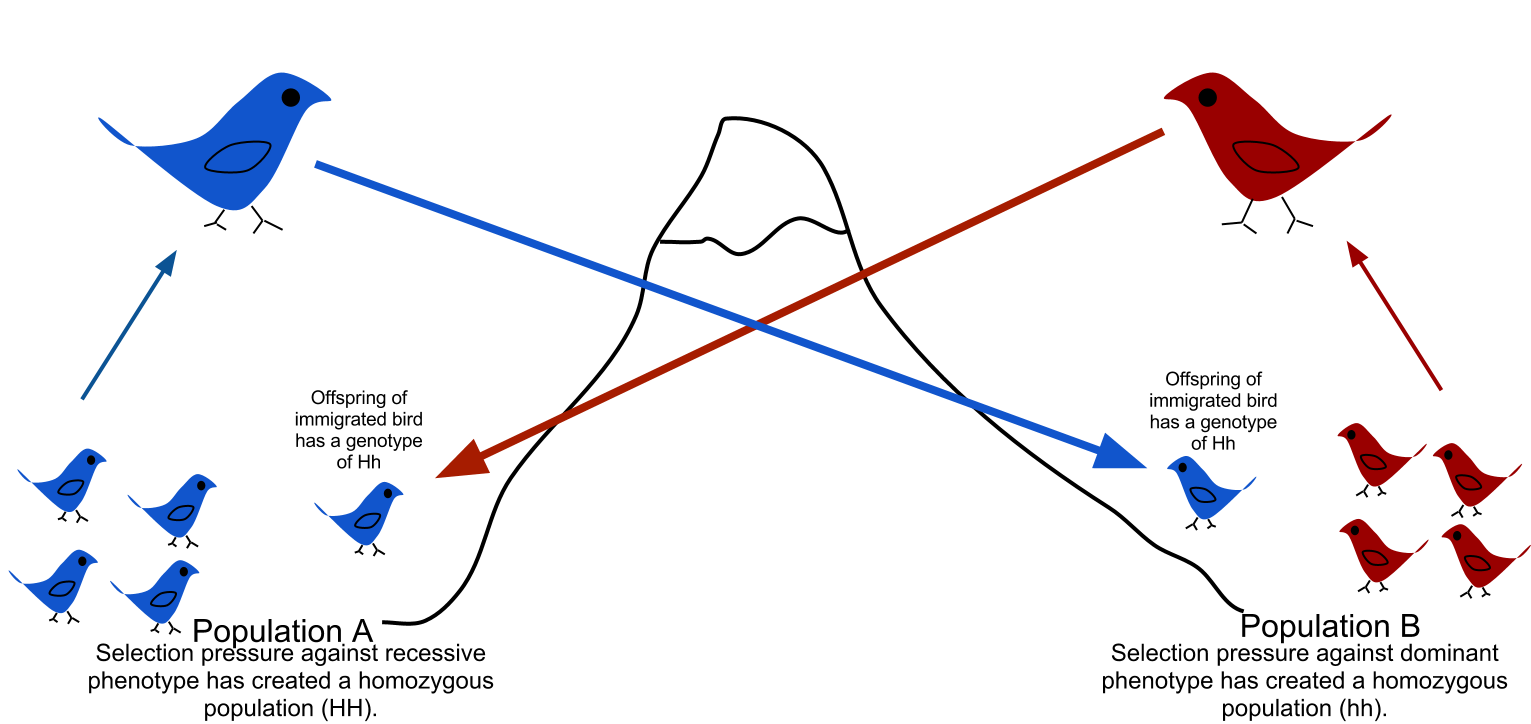
In population genetics, gene flow (also known as migration and allele flow) is the transfer of genetic material from one population to another. If the rate of gene flow is high enough, then two populations will have equivalent allele frequencies and therefore can be considered a single effective population. It has been shown that it takes only "one migrant per generation" to prevent populations from diverging due to drift.[1] Populations can diverge due to selection even when they are exchanging alleles, if the selection pressure is strong enough.[2][3] Gene flow is an important mechanism for transferring genetic diversity among populations. Migrants change the distribution of genetic diversity among populations, by modifying allele frequencies (the proportion of members carrying a particular variant of a gene). High rates of gene flow can reduce the genetic differentiation between the two groups, increasing homogeneity.[4] For this reason, gene flow has been thought to constrain speciation and prevent range expansion by combining the gene pools of the groups, thus preventing the development of differences in genetic variation that would have led to differentiation and adaptation.[5] In some cases dispersal resulting in gene flow may also result in the addition of novel genetic variants under positive selection to the gene pool of a species or population (adaptive introgression.[6])
There are a number of factors that affect the rate of gene flow between different populations. Gene flow is expected to be lower in species that have low dispersal or mobility, that occur in fragmented habitats, where there is long distances between populations, and when there are small population sizes.[7][8] Mobility plays an important role in dispersal rate, as highly mobile individuals tend to have greater movement prospects.[9] Although animals are thought to be more mobile than plants, pollen and seeds may be carried great distances by animals, water or wind. When gene flow is impeded, there can be an increase in inbreeding, measured by the inbreeding coefficient (F) within a population. For example, many island populations have low rates of gene flow due to geographic isolation and small population sizes. The Black Footed Rock Wallaby has several inbred populations that live on various islands off the coast of Australia. The population is so strongly isolated that lack of gene flow has led to high rates of inbreeding.[10]
Measuring gene flow[edit]
The level of gene flow among populations can be estimated by observing the dispersal of individuals and recording their reproductive success.[4][11] This direct method is only suitable for some types of organisms, more often indirect methods are used that infer gene flow by comparing allele frequencies among population samples.[1][4] The more genetically differentiated two populations are, the lower the estimate of gene flow, because gene flow has a homogenizing effect. Isolation of populations leads to divergence due to drift, while migration reduces divergence. Gene flow can be measured by using the effective population size () and the net migration rate per generation (m). Using the approximation based on the Island model, the effect of migration can be calculated for a population in terms of the degree of genetic differentiation().[12] This formula accounts for the proportion of total molecular marker variation among populations, averaged over loci.[13] When there is one migrant per generation, the inbreeding coefficient () equals 0.2. However, when there is less than 1 migrant per generation (no migration), the inbreeding coefficient rises rapidly resulting in fixation and complete divergence ( = 1). The most common is < 0.25. This means there is some migration happening. Measures of population structure range from 0 to 1. When gene flow occurs via migration the deleterious effects of inbreeding can be ameliorated.[1]
The formula can be modified to solve for the migration rate when is known: , Nm = number of migrants.[1]




Human assisted gene-flow[edit]
Genetic rescue[edit]
Gene flow can also be used to assist species which are threatened with extinction. When a species exist in small populations there is an increased risk of inbreeding and greater susceptibility to loss of diversity due to drift. These populations can benefit greatly from the introduction of unrelated individuals[11] who can increase diversity[16] and reduce the amount of inbreeding, and potentially increase population size.[17] This was demonstrated in the lab with two bottleneck strains of Drosophila melanogaster, in which crosses between the two populations reversed the effects of inbreeding and led to greater chances of survival in not only one generation but two.[18]
While gene flow can greatly enhance the fitness of a population, it can also have negative consequences depending on the population and the environment in which they reside. The effects of gene flow are context-dependent.