Ion channel
Ion channels are pore-forming membrane proteins that allow ions to pass through the channel pore. Their functions include establishing a resting membrane potential,[1] shaping action potentials and other electrical signals by gating the flow of ions across the cell membrane, controlling the flow of ions across secretory and epithelial cells, and regulating cell volume. Ion channels are present in the membranes of all cells.[2][3] Ion channels are one of the two classes of ionophoric proteins, the other being ion transporters.[4]
Not to be confused with Ion Television or Ion implantation.The study of ion channels often involves biophysics, electrophysiology, and pharmacology, while using techniques including voltage clamp, patch clamp, immunohistochemistry, X-ray crystallography, fluoroscopy, and RT-PCR. Their classification as molecules is referred to as channelomics.
Biological role[edit]
Because channels underlie the nerve impulse and because "transmitter-activated" channels mediate conduction across the synapses, channels are especially prominent components of the nervous system. Indeed, numerous toxins that organisms have evolved for shutting down the nervous systems of predators and prey (e.g., the venoms produced by spiders, scorpions, snakes, fish, bees, sea snails, and others) work by modulating ion channel conductance and/or kinetics. In addition, ion channels are key components in a wide variety of biological processes that involve rapid changes in cells, such as cardiac, skeletal, and smooth muscle contraction, epithelial transport of nutrients and ions, T-cell activation, and pancreatic beta-cell insulin release. In the search for new drugs, ion channels are a frequent target.[8][9][10]
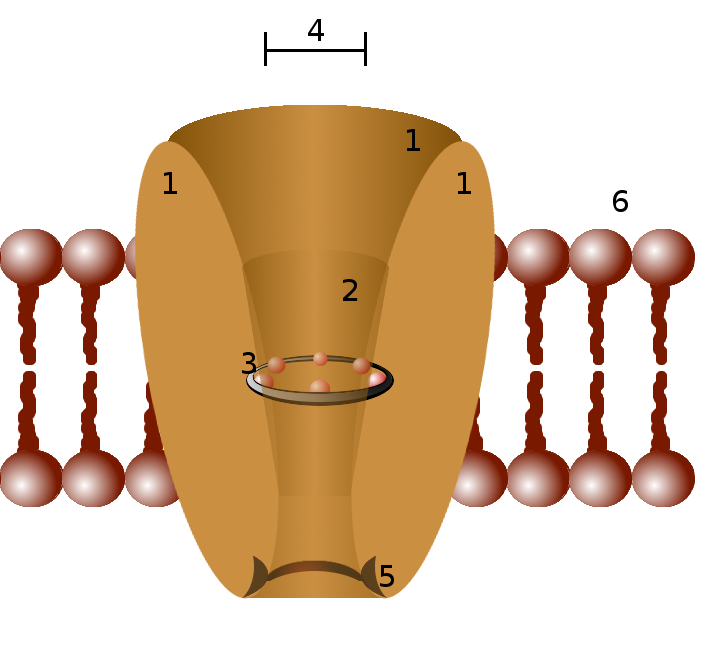
Detailed structure[edit]
Channels differ with respect to the ion they let pass (for example, Na+, K+, Cl−), the ways in which they may be regulated, the number of subunits of which they are composed and other aspects of structure.[29] Channels belonging to the largest class, which includes the voltage-gated channels that underlie the nerve impulse, consists of four or sometimes five [30] subunits with six transmembrane helices each. On activation, these helices move about and open the pore. Two of these six helices are separated by a loop that lines the pore and is the primary determinant of ion selectivity and conductance in this channel class and some others. The existence and mechanism for ion selectivity was first postulated in the late 1960s by Bertil Hille and Clay Armstrong.[31][32][33][34][35] The idea of the ionic selectivity for potassium channels was that the carbonyl oxygens of the protein backbones of the "selectivity filter" (named by Bertil Hille) could efficiently replace the water molecules that normally shield potassium ions, but that sodium ions were smaller and cannot be completely dehydrated to allow such shielding, and therefore could not pass through. This mechanism was finally confirmed when the first structure of an ion channel was elucidated. A bacterial potassium channel KcsA, consisting of just the selectivity filter, "P" loop, and two transmembrane helices was used as a model to study the permeability and the selectivity of ion channels in the Mackinnon lab. The determination of the molecular structure of KcsA by Roderick MacKinnon using X-ray crystallography won a share of the 2003 Nobel Prize in Chemistry.[36]
Because of their small size and the difficulty of crystallizing integral membrane proteins for X-ray analysis, it is only very recently that scientists have been able to directly examine what channels "look like." Particularly in cases where the crystallography required removing channels from their membranes with detergent, many researchers regard images that have been obtained as tentative. An example is the long-awaited crystal structure of a voltage-gated potassium channel, which was reported in May 2003.[37][38] One inevitable ambiguity about these structures relates to the strong evidence that channels change conformation as they operate (they open and close, for example), such that the structure in the crystal could represent any one of these operational states. Most of what researchers have deduced about channel operation so far they have established through electrophysiology, biochemistry, gene sequence comparison and mutagenesis.
Channels can have single (CLICs) to multiple transmembrane (K channels, P2X receptors, Na channels) domains which span plasma membrane to form pores. Pore can determine the selectivity of the channel. Gate can be formed either inside or outside the pore region.
There are a number of disorders which disrupt normal functioning of ion channels and have disastrous consequences for the organism. Genetic and autoimmune disorders of ion channels and their modifiers are known as channelopathies. See Category:Channelopathies for a full list.
History[edit]
The fundamental properties of currents mediated by ion channels were analyzed by the British biophysicists Alan Hodgkin and Andrew Huxley as part of their Nobel Prize-winning research on the action potential, published in 1952. They built on the work of other physiologists, such as Cole and Baker's research into voltage-gated membrane pores from 1941.[41][42] The existence of ion channels was confirmed in the 1970s by Bernard Katz and Ricardo Miledi using noise analysis . It was then shown more directly with an electrical recording technique known as the "patch clamp", which led to a Nobel Prize to Erwin Neher and Bert Sakmann, the technique's inventors. Hundreds if not thousands of researchers continue to pursue a more detailed understanding of how these proteins work. In recent years the development of automated patch clamp devices helped to increase significantly the throughput in ion channel screening.
The Nobel Prize in Chemistry for 2003 was awarded to Roderick MacKinnon for his studies on the physico-chemical properties of ion channel structure and function, including x-ray crystallographic structure studies.